

The electrons in a wire with no Electric Field are generally moving around in a random fashion.
When
an Electric Field is applied to the ends of the wire
(a battery is hooked-up) there is a
net movement of electrons along the Electric Field.(in
one direction)
The velocity of the electrons
after the Electric Field is applied is actually very
slow.
The Electric Field is fast and is applied throughout
the wire almost instantaneously.



(Watts = Joules/Second)


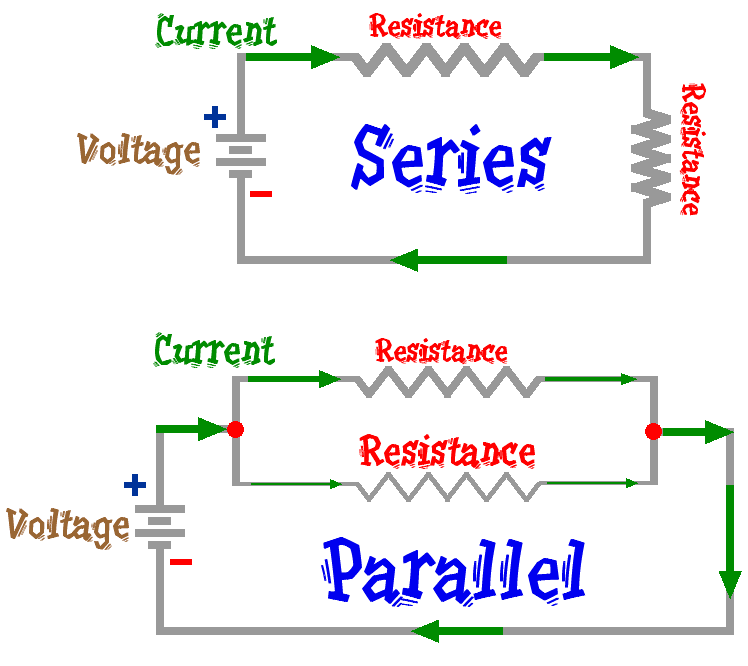




The Total Resistance in the circuit RT = R1 + R2 + . . .
The Current is the same thru every resistor in the circuit
The Total Voltage in the circuit VT = V1 + V2 + . . .
In a Parallel Component in a Circuit
The Total Resistance in
the component 1/RT = 1/R1
+ 1/R2 + . . .
The Total Current in the
component IT = I1
+ I2 + . . .
The Voltage over each Resistor
in each branch of the component
is the same.

Capacitance Circuits
Charge is released from the capacitor in a certain amount of time = RC

Basic Electrical Safety
Heartpacemakers
The physiological effect of current
Basic Electrical Safety
* To flow electricity must have a complete path.
* Electricity flows through conductors
* water, metal, the human body
* The human body is a conductor
* More than 3 ma
* painful shock* More than 10 ma
* muscle contraction "no-let-go" danger
* More than 30 ma
* lung paralysis- usually temporary
* More than 50 ma
* possible ventricular fob. (heart dysfunction, usually fatal)
* 100 ma to 4 amps
* certain ventricular fibrillation, fatal
* Over 4 amps
* heart paralysis; severe burns. Usually caused by >600 volts
The
majority of all action potentials are generated in the axon hillock. However
in sensory neurons the action potential is generated by the peripheral
(axonal) process, just proximal to the receptor region. These areas are
also known as the trigger regions.
An action potential is
generated due to membrane potential reaching threshold due to a graded
potential. Threshold is a membrane
potential at which the membrane in the trigger region reaches approximately
-55mV, a depolarization of about 15 mV.
At
this point action potentials become self propagating. This
means that one action potential automatically triggers the neghboring
membrane areas into producing an action potential. Thus once threshold
is reached action potentials always propagate down the axon to the synaptic
or secretory regions of the axon.
The actual process of the action potential
generation occurs in four steps, consecutive,
but overlapping. These steps are all opening and/or closing of ion gates,
and subsequent changes in membrane potentials. Read
on. . . .
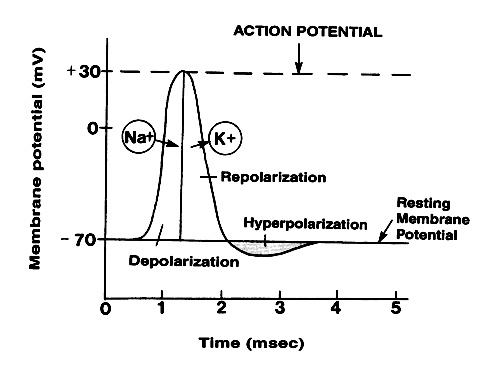
1) The first step is the resting state, where all active ion channels are closed. Almost all voltage gated sodium and potassium gates are closed. However some potassium is leaking out via leakage channels, and even smaller amounts of sodium are diffusing in.
2a) This phase is actually consists of two substeps. As the trigger region membrane is depolarized to threshold voltage gated sodium channels begin to open. By the time threshold potential is reached enough voltage gated sodium channels (vgsc's) are opened that the potential is now self generating, being driven on by the influx of Na+. With the vast majority of the vgsc's opened Na+ floods into the cell, further depolarizing the cell, and increasing the membranes permeabliliy to sodium by over 1000 times. Eventually the cell lets in so many positively charged sodium ions that the membrane potential goes from -70mV to +30mV.
2b) As the membrane potential reaches 0 mV, and the cell interior becomes more and more positive, sodium entry becomes less rapid, as the electrical gradient starts to repel the ions. Furthermore in less than a milisecond of reaching threshold the sodium gates begin to close, albeit slowly. This additionally causes the membrane to start to loose permeablility with regard to the sodium ions. As the net influx of sodium declines, and then finally stops, the membrane has reached it's maximum depolarization at about +30mV.
3) As the membrane potenial approachs +30 mV, voltage gated potassium channels open and positively charged potassium ions begin to flow out of the cell. This begins to repolarize, the cell by reducing the excess internal positive charge and moving the membrane potential closer to the resting potential. At this point the cell is basically impermeable to sodium and very permeable to potassium which rapidly flows out of the cell down both it's electrical (initially) and chemical gradients.
4) Potassium efflux (exiting) continues past the resting potential of -70 mV due to the slow closing voltage gated potassium channels. This causes a hyperpolarization know as undershoot which takes the membrane potential to around -75mV. Soon afterward the cell returns to resting potential via the standard membrane proteins.
Read on. . . .
PBS Neurons
Holy Animations, Batman!
An action potential
Action Potential II
Action Potential III
Action Potential Theory
Physics Today May 1948
Arthur K. Solomon
In May 1948, Arthur Kaskel Solomon,
an associate professor of physiological chemistry at Harvard University,
offered a fresh perspective on the relation between biology and physics--the
field of his own PhD--and gave a glimpse into how the mysteries of cancer
were viewed those 54 years ago. He retired as a professor of biophysics
from Harvard Medical School in 1983.
The connection between physics and cancer is peculiarly
intimate--as intimate as cause and effect. For radiation, a valuable
agent in the treatment of cancer, can itself cause cancer.
This curious interrelationship exists largely
because no one knows what causes growth, whether it be normal or abnormal.
The primary problem in cancer is not an exploration of such isolated problems
as the connection between radiation and cancer. It is the much larger
and much more stimulating problem of understanding the fundamentals of
growth.
The first necessary step, therefore, is an examination
of the contributions that physics can make to the larger problem of growth,
rather than to the more limited and purely applied problem of the treatment
of cancer. This is just another way of asking the even more fundamental
question: What can physics bring to
biology? Precisely because cancer is not yet understood,
and precisely because the explanation may lie in any of the areas of biological
research, the attack on the problem demands a wide expansion in our understanding
of the fundamental processes of biology.
What can physics bring to biology? The answer
to the problem is subjective, and the answer of a physicist will probably
be quite different from that of a biologist. At the outset, the biologist
wants techniques; the physicist believes that his greatest contribution
is conceptual. And the truth of the matter, no doubt, lies in that broad
no man's land between these extremes.
Techniques of physics
Today, the use of radioactive isotopes is a very fashionable
technique. Partly this is the inevitable result of the terrible fascination
of the atom bomb, and partly it is the result of the real extension of
basic biological knowledge that has already come about from the use of
isotopes. Using radioactive isotopes, certain compounds can be traced
without minute control of the experimental biological system. The final
fate of an essential foodstuff may be determined by labeling it with radioactive
atoms, then finding how these are excreted during the animal's lifetime
or, after post-mortem, where they have concentrated. A principle such
as this can be extended to smaller and smaller systems and leads to an
intimate knowledge of some of the basic chemical systems that exist in
living matter. It is even possible to make experiments on normal human
adults, if the measurements are confined to excretory products and blood.
Indeed, some students at Harvard Medical School are earning a small stipend
by acting as experimental animals in studies of the uptake of minute quantities
of radioactive iron in normal blood.
The tracer technique
is applicable to studies other than the unraveling of biochemical reactions.
Wherever a fluid diffuses through a barrier, wherever a fluid is transmitted
through a hollow vessel, radioactive tracers make it possible to measure
the diffusion and the flow. And wherever small amounts of material, too
small to be detected by normal chemical means, play an important role
in a physiological process, tracers help elucidate the pathways.
These successful physical and chemical excursions
into the domain of the microcosmos bring to mind that early great contribution
and introduction into the microworld that came to biology by way of physics
through the lens, the telescope, and the microscope. Physics, pure and
applied, has continued to contribute to the development of the microscope
from that first incredibly crude instrument used to such good purpose
by Leuwenhoek. Today the polarizing microscope and the phase microscope
represent the newest development of these instruments routinely available
for the use of biologists. To progress even further into the microworld,
one must turn to the ultraviolet microscope, the electron microscope,
and even to the proton microscope now being developed in France.
The ultimate limit of vision with a conventional
microscope is the wavelength of the light with which the image is viewed.
Therefore, the shorter the wavelength, the tinier the object that can
be discerned. Since the ultraviolet microscope uses shorter waves than
the visible, a consequent decrease in the lower visual limit may be expected.
In modern physics, all particles have characteristic wavelengths, the
heavier and faster the particle the shorter the wave. But even a particle
as light as an electron has a characteristic wavelength many times shorter
than the wavelength of light. Thus the electron microscope can 'see' objects
far smaller than those visible in the best conventional microscope. It
may be possible to see even smaller objects by the use of heavier particles
at the same speed, as, for example, the proton, 1800 times heavier than
the electron. If the technical difficulties involved in the construction
of a proton microscope can be solved, we may penetrate further into ultimate
structure.
Variables and more variables
It is always easier to talk about the specific
contributions that techniques make to a sister science than to explain
the conceptual differences between sciences. This is particularly true
since one man's opinion can never truly represent the whole profession.
In general, the approach of the physicist is simple and direct
because his problems can usually be set up in simple and direct terms.
In the main, his variables are controllable. But coupled
with the dry precision of the experimental approach are the vast imaginative
powers of theory. The first floor of
the physical structure is built on the solid foundation of experimental
fact; the lofty towering spire comes directly from the fancy of theory.
Theory must be mathematically developed and exactly stated, and it remains
tenable only so long as no new experiment comes to tumble the structure
down again.
In sharp contradistinction to physics, all biological
processes are very dependent on a large number of interrelated variables.
The simplest sample to be studied is that smallest fraction of living
tissue which will still carry out the functions that are being investigated.
An attempt to apply too stringent a control of the variables may mean
destruction of the experimental system. Many reactions that can be carried
out in the normal animal liver cannot be reproduced if the liver is ground
up and placed in a glass vessel to provide an environment and a temperature
that can be accurately controlled.
In the past, the biologist has never
failed to produce theories to account for his experimental results; the
theories, however, can rarely be developed in exact mathematical terms.
This is particularly true because so few biological experiments can be
set up so that the variables can be controlled at the will of the investigator.
The complexity of biology is, indeed, the complexity of life, presenting
a multitude of problems that can barely be attacked at all. Such complexity
does not yield readily to mathematical treatment. Our understanding of
biology rests on a knowledge of the basic chemistry and physics of biological
systems. Surely it is logical to call in the physicist and chemist themselves
in a joint attack on the problem of growth, so that we can deploy the
best talents available for the steps ahead. Perhaps the greatest of their
contributions will be the concept and discipline of these more exact sciences.
Tools
for treatment
(My research at SDSU)
In the light of the larger problems of growth,
the physicist's contribution to the study of cancer has been modest. The
present and immediate contributions are those that physics can make to
the healing of disease, and these are now, as they have been in the past,
very limited. The treatment of sick people does not, in general, lead
to a knowledge of what made them sick.
The present-day use of radiation therapy is merely
an extension of the familiar use of radium and x rays in the treatment
of cancer.  The
action of radiation in the body is not completely understood, but there
is reasonable agreement that it is the effect of ionization (in which
atoms or molecules acquire an electric charge) along the path of radiation.
Ionization by x rays cannot be localized at one specific point along the
path of the x ray in tissue: Thus, treatment of deep-seated tumors with
x rays irradiates not only the tumor but also the skin and intervening
tissues. A more specific method of introducing the radiation directly
at the tumor would be a great help.
The
action of radiation in the body is not completely understood, but there
is reasonable agreement that it is the effect of ionization (in which
atoms or molecules acquire an electric charge) along the path of radiation.
Ionization by x rays cannot be localized at one specific point along the
path of the x ray in tissue: Thus, treatment of deep-seated tumors with
x rays irradiates not only the tumor but also the skin and intervening
tissues. A more specific method of introducing the radiation directly
at the tumor would be a great help.
It now seems clear that such an effect can be
achieved by using electrons as irradiating agents. The electron cannot
penetrate tissue to the depth reached by x-rays, but the ionization is
much more intense, and increases just at the end of the electron path.
The great energy required to propel the electrons deep into tissue has
only recently become available with the development of the synchrotron,
a tool in nuclear physics devised to provide an intense beam of high-energy
electrons. Its use in biology stems from the ability of these
electrons to produce a local region of dense ionization deep inside the
body with very little exposure at the skin. A good deal
of research is needed to show just how effective such dense, localized
ionization can be: research not only in the physical, but also in the
clinical sense, since it is often impossible to know where the radiation
should be directed even when it can be controlled.
The long way
The physicist and the doctor both tend to be cautious
now in predicting the future effect of newly developed tools, because
of their joint experience with radioisotope therapy. In the early days
before the war, it was thrilling to devote long cyclotron hours to the
bombardment of phosphorus for the treatment of a leukemic patient. Early
results were promising: The radioactive phosphorus seemed to concentrate
in the bone marrow, where white blood cells are made. Since leukemia is
characterized by the overproduction of white blood cells, it was sensible
to hope that radiophosphorus could deliver enough radiation in the right
places to control this overproduction. Although control of this sort was
exercised, our initial optimism has given place to the realization that,
in general, the average prolongation of life of leukemic patients is no
greater with radiophosphorus than with the more conventional x-ray treatment.
Some initial striking success has been obtained
with the use of radioactive iodine in the treatment of thyroid cancer.
Since the thyroid picks up iodine preferentially, it seemed, by the same
rationale that applied to phosphorus treatment, that thyroid cancer might
be controlled by delivering the iodine radiation directly into the thyroid
itself. But not all thyroid cancers take up iodine; indeed, the more vicious
the cancer, the less likely it is to accumulate iodine. Nonetheless, a
few cases have been controlled by iodine treatment, particularly patients
who have had surgical thyroid removal, and who are still troubled by malignant
transplants of small fractions of thyroid tissue at different places throughout
the body. Such small amounts of malignant tissue have, in one case at
least, been successfully controlled for a period of years. Thyroid cancer,
however, is a relatively rare form of cancer, and the percentage of thyroid
patients who respond to radioactive treatment is small. To sum up: The
use of radioactive isotopes in the treatment of cancer has not yet proved
itself on an appreciable scale, despite repeated efforts.
One other fashion in which radioactive isotopes
may produce a great impact on cancer is in diagnosis. It has already been
shown that breast cancers pick up appreciably more radioactive phosphorus
than do surrounding sections of normal tissue, presumably because the
tumorous tissue is growing so much faster than normal that it incorporates
all of its structural elements very rapidly. If this abnormality in phosphorus
uptake can be used to diagnose and to locate not only breast cancers,
but other and more deep-seated lesions, then, indeed, it will have proved
itself of great value in the treatment of cancer.
Until that time, realistic appraisal of the cancer problem brings us back
to our starting point: The
real contribution of physics to cancer lies in the joint prosecution,
by the physicist, by the chemist, by the biologist, of basic research
on the problems of growth.